РЕГИОНАЛЬНЫЙ ИНФОРМАЦИОННЫЙ ДАЙДЖЕСТЭКОНОМИКА, ЭКОЛОГИЯ, ИСТОРИЯ, КУЛЬТУРА |

Первая общественная российско-американская конференция
«Проблемы сохранения биоресурсов Берингова моря»
Современное состояние компонентов экосистемы Берингова моря
Дулепова Е.П.
вед. научн. сотр. лаб. прикладной биоценологии
ТИНРО-центра,
Владивосток, тупик Шевченко, 4
Для того чтобы судить о состоянии и функционировании экосистемы, необходимы точные знания как о современном статусе ее компонентов, так и об основополагающих (главным образом трофических) связях между ними.
С начала 1980-х гг. НТО ТИНРО в Беринговом море широким фронтом ведутся крупномасштабные комплексные исследования, при этом ключевая роль принадлежит количественной оценке обилия, межгодовой динамике и продуктивности основных компонентов экосистемы. Объем информации, полученной за весьма длительный период исследований, позволяет представить целостную картину состояния биоты этого водоема и направленность межгодовой динамики основных ее составляющих. Немаловажная роль в этих комплексных исследованиях в последнее время принадлежит оценке низших трофических уровней (фитопланктону, бактериопланктону, простейшим и зоопланктону). При этом работы в данном направлении ведутся не только силами ТИНРО, но и Академии наук. В настоящее время В.П. Шунтовым закончена крупная работа, посвященная состоянию, динамике и оценке указанных компонентов в дальневосточных морях. Современные оценки биомасс и продукции всех групп гидробионтов, формирующих низшие трофические уровни (от фитопланктона и бактерий до зообентоса), находятся на более высоком уровне, чем это представлялось ранее (табл. 1). С такими оценками в большей степени согласуются современные количественные оценки представителей высших трофических уровней, в том числе тех, кто формирует биологические ресурсы и сырьевую базу рыболовства.
Таблица 1. Ранжирование дальневосточных морей и сопредельных районов северо-западной части Тихого океана по обилию биогенов, биомассам и продукции основных групп гидробионтов (Шунтов, 2000, с дополнениями и изменениями)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечание: К. — курильские воды океана, К.К. — камчатско-командорские воды океана, Б.м. — Берингово море, О.м. — Охотское море, Я.м. — Японское море.
Лейтмотивом настоящей конференции стали тревожные выводы о кризисной ситуации в экосистеме Берингова моря. При этом имеются в виду некоторые следствия, связываемые с аномальным потеплением и антропогенным воздействием на природу морей: обилие кокколитофорид, повышенная смертность птиц и морских млекопитающих, снижение количества минтая и других рыб.
С экосистемных позиций под кризисной ситуацией в экосистеме подразумевается нарушение ее функционирования, выражающееся не только в падении численности тех или иных видов и групп, но и в необратимом нарушении структурных связей.
Важным итогом биоценологических исследований является получение современных данных о суммарном запасе и продукции зоопланктона. В отношении общих биомасс зоопланктона не было замечено до 1995 г. какой-либо устойчивой тенденции долгопериодной изменчивости. Сравнение данных по биомассе зоопланктона в Анадырско-Наваринском районе за 1980-е гг. и 1998 г. показало, что биомасса основных и важных с кормовой точки зрения планктонных групп даже возросла (рис.1). В то же время в отдельных районах отмечается существенная межгодовая изменчивость. В Анадырском заливе осенью 1998 г. биомасса возросла многократно. В следующем 1999 г., который оказался более холодным, биомасса зоопланктона вновь снизилась. Особенно мало планктона было в начале лета. Но в конце лета и осенью биомасса зоопланктона лишь в два раза была ниже, чем в 1998 г. И связано это было в основном с уменьшением количества эвфаузиид (однако порядок цифр в данной ситуации все равно достаточно высок). В 2000 г., который оказался еще холоднее, чем 1999 г., биомасса планктона в Анадырском заливе еще более уменьшилась, и в то же время в Наваринском районе — увеличилась. Таким образом, изменения в общей биомассе зоопланктона, скорее всего, вписываются в картину межгодовой динамики этой характеристики.
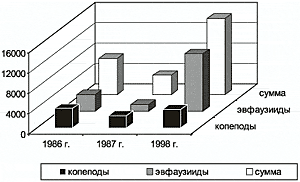
Рис. 1. Динамика биомассы (тыс. т) копепод и эвфаузиид в Анадырско-Наваринском районе Берингова моря.
В то же время при снижении количества минтая, нагуливающегося в этом районе, годовое потребление пищи осталось приблизительно на одном уровне — 15,5 млн т в 1980-х гг. и 14 млн т в 1998 г. — в связи с тем, что интенсивность питания возросла. Это свидетельствует о том, что в самой трофической структуре заложен порядочный задел прочности.
В 1990-е гг. по различным районам дальневосточных морей, включая Берингово, прошла волна повышенной численности хищного зоопланктона. Если в 1980-е гг. на долю хищного зоопланктона в юго-западной части Берингова моря приходилось около 27 % от общей биомассы, то в 1991 г. на его долю здесь приходилось 54 %, в 1993 г. — 57 %. Сильное увеличение доли хищников, по-видимому, инициировано значительным снижением биомассы такого планктоноядного вида, как минтай, который может рассматриваться как конкурент хищного зоопланктона за пищу. При этом сильное увеличение доли хищников в зоопланктоне является таким уровнем, когда в результате авторегуляции был неизбежен возврат к исходному состоянию. И действительно, в 1995 г. доля хищного зоопланктона уменьшилась в юго-западной части до 34 %. В последующие годы снизилась еще более существенно и варьировала по районам в 1998 г. — от 22 до 39 %, а в 1999 г. — от 16 до 31 % (рис. 2).
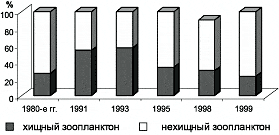
Рис. 2. Динамика доли хищного зоопланктона в юго-западной части Берингова моря.
В целом в ряду крупных районов российских экономических вод наиболее высокие суммарные биомассы и продукция гидробионтов высших трофических уровней наблюдается именно в Беринговом море, где ниже средние показатели, характеризующие низшие трофические уровни. В генерализованном виде оценки нектона и нектобентоса выглядят следующим образом. С учетом данных по восточной части моря, биомасса рыб для всего моря в 1980-е гг. была оценена в 60–50 млн т при годовой продукции 23 млн т (минтай 20–25 млн т, мезопелагические рыбы — 15–20, донные — 11, прочие пелагические — 5), кальмаров — 4 млн т, крабов — 1,2 млн т, креветок — 1 млн т, букцинид — 0,25 млн т. На российскую экономическую зону здесь приходилось всего 12,5–15 млн т рыб (минтай — 5,5–6,5, донные — 2,2, мезопелагические — 3 млн т, прочие пелагические — 1,5).
В начале 1990-х гг. были зафиксированы перестройки в рыбных сообществах пелагиали всех дальневосточных морей, в частности выразившиеся в снижении численности такого вида, как минтай (и на юге — иваси). Снижение эффективности воспроизводства и количества минтая с конца 1980-х гг., начавшись в зал. Аляска и восточной части Берингова моря, перешло в его западную часть и затем в Охотское море. Запас беринговоморского минтая, воспроизводящегося в основном в Олюторском и Карагинском заливах Берингова моря, последовательно снижался с начала 1990-х гг. Но уменьшение численности минтая сопровождалось возрастанием ресурсов корфо-карагинской сельди, что свидетельствует о смене доминирующего вида в пелагическом ихтиоцене западной части Берингова моря.
Современный пониженный уровень рыбопродуктивности дальневосточных морей и Берингова моря — это естественный процесс и не является неожиданным, а, кроме того, был предсказан со значительной заблаговременностью (Шунтов, 1986; Шунтов и др., 1997). Минтай, так же как и сардина-иваси, принадлежит к относительно теплолюбивым видам, численности которых свойственна долгопериодная цикличность. Промысел и, соответственно, изучение минтая в северо-западной части Тихого океана пока охватили только один цикл динамики его запасов, да и то не полностью. Точный уровень его биомассы до начала масштабного промысла неизвестен. Ее рост в начале 1970-х гг. был определен появлением ряда урожайных поколений во второй половине 1960-х гг. В климато-океанологическом окружении и изменчивости биологической структуры макроэкосистем особая важность придается 40–60-летней цикличности этих позиций, и этот период пониженной численности минтая продолжится еще, по крайней мере, два десятилетия (Шунтов, 1999, 2000; Radchenko, 2001). А так как в это время пониженный запас будет иметь целый ряд важных промысловых объектов, то оценки (при формальном подходе, что нередко имеет место сейчас) о кризисном состоянии экосистемы будут неизбежны и в дальнейшем.
Механизм формирования урожайности поколений промысловых объектов — процесс сложный и многофакторный. Многочисленные факторы, оказывающие влияние на численность, с известной долей условности можно подразделить на несколько уровней: 1 — космофизические и глобальные климатоокеанологические факторы, формирующие крупные периоды или эпохи; 2 — климатические и гидрологические условия конкретного года и конкретного района; 3 — биоценологические отношения, связанные с фактором плотности (в первую очередь конкуренция за пищу); 4 — хищничество на разных стадиях онтогенеза; 5 — популяционные факторы, в том числе авторегуляционного характера; 6 — антропогенное влияние на биоту и ее окружение, в том числе промысел, аквакультура, загрязнение. Из-за трудности учета всех факторов, их значимости, а также при наличии вариаций при их комбинированном действии теоретическое предсказание тенденций динамики численности очень затруднено, а зачастую реальный ход событий оказывается непредсказуем. Вот поэтому в практике рыбохозяйственного прогнозирования исключительное значение придается регулярному мониторингу за состоянием единиц запасов и оценке эффективности воспроизводства и учету ближнего пополнения промыслового запаса.
Донные сообщества, как известно, по сравнению с пелагическими более консервативны и стабильны. Тем не менее и в них происходят определенные изменения. Для оценки современной ситуации в нектонных и донных сообществах северо-западной части Берингова моря в конце 1990-х гг. ТИНРО-центром был проведен целый ряд комплексных экспедиций. Сопоставляя современный состав траловых уловов на мелководье северо-западной части Берингова моря с аналогичными данными за 1985 г., нельзя не заметить их принципиальное отличие. Если раньше в уловах явно преобладала треска, доля которой в отдельных районах достигала 80 %, то сейчас наблюдается иная картина. В Олюторском и Анадырском заливах видовая структура уловов стала более разнообразной, и лишь на корякском шельфе треска сохранила свое прежнее лидирующее положение среди донных рыб, но и здесь ее вклад снизился с 75,3 до 55,1 %. Однако общая биомасса донных рыб по сравнению с прошлыми годами в 2000 г. значительно выше. Если в этот год в Олюторском заливе в пределах глубины 50–300 м она составила 130,74 тыс. т, то в 1998 г., включая и часть Карагинского залива с материковым склоном, — 63,15 тыс. т, а в 1999 г. — всего около 6 тыс. т, что в несколько раз меньше со сравниваемыми годами.
Появившиеся в последнее время тревожные заключения о кризисной ситуации в Беринговом море на низших трофических уровнях, связанные с загрязнением и аномальным потеплением, противоречат реальной картине. Отдельные локальные прибрежные районы дальневосточных морей (например зал. Петра Великого) уже сейчас сильно загрязнены и их экологическое состояние может трактоваться как кризисное. Но, к счастью, дальневосточные моря в целом сейчас являются чистыми и загрязнение пока не является здесь фактором, нарушающим нормальное функционирование макроэкосистем и воспроизводство гидробионтов. Что касается влияния глобального потепления, то сейчас нет никаких оснований ставить эту проблему в разряд приоритетных. По крайней мере, в течение нескольких десятков лет. Кроме того, годы на рубеже последних столетий оказались в дальневосточных морях аномально холодными. И в то же время следует заметить, что при потеплении рыбопродуктивность наших морей будет выше.
В контексте изложенного представляются неубедительными диагнозы состояния экосистем Берингова и Чукотского морей по данным работ по программе БЕРПАК. После первых экспедиций был сделан вывод о том, что экосистема Берингова моря не подвержена антропогенному воздействию и что за 30–50 лет в ней не произошли какие-либо существенные изменения (Всесторонний анализ экосистемы Берингова моря, 1987). Это писалось о море, где к тому времени были основательно выбиты представители высших трофических уровней — киты и тюлени, а также допущен перелов мощных стад камбал, окуней и палтусов. Через некоторое время лейтмотив выводов по итогам программы БЕРПАК совершенно изменился, при этом особое значение придается экологическим последствиям за-грязнения морей (в том числе эвтрофикации вод под влиянием антропогенных факторов), парникового эффекта, истощения озонного слоя и увеличения ультрафиолетовой радиации. А в подтверждение этого тревожного диагноза говорится о снижении видового разнообразия зоопланктона (Динамика экосистемы Берингова и Чукотского морей, 2000). Ни один из этих драматических выводов цитируемой монографии не подтвержден убедительными доказательствами. Что же касается реального состояния компонентов, формирующих низшие трофические уровни экосистемы Берингова моря, то они находятся в нормальном функциональном состоянии. Берингово море как было, так и остается одним из весьма биопродуктивных морей Мирового океана. Как азиатские, так и американские его берега и сопредельные районы материков в основном пока не охвачены крупномасштабной хозяйственной деятельностью, они слабо заселены, здесь нет крупных портов и городов. Это чистое море, а наблюдаемые изменения в структуре и количественных характеристиках компонентов, слагающих в биоценозах низшие трофические уровни, находятся в пределах свойственной естественной межгодовой и многолетней динамики. Этот вывод за некоторыми локальными исключениями может быть перенесен на всю акваторию дальневосточных российских вод.
В истории промысла в дальневосточных морях известно много случаев перепромысла, касающихся в первую очередь донных гидробионтов (крабы, камбалы, палтусы) и представителей высших трофических уровней (киты, тюлени и даже морские птицы). В преобладающем большинстве случаев при внедрении в практику регулирующих и охранных мер имело и имеет место восстановление биомасс и численности, о чем свидетельствует весьма объемный современный промысел и регулярный мониторинг биоресурсов. В ряду целого ряда таких видов пока не поддается объяснению лишь современное снижение количества сивучей и калана, которые уже много десятилетий не являются объектами промысла. Что касается флюктуирующих видов, то современный пониженный уровень численности некоторых из них не выходит за пределы известных циклических колебаний их количества.
Изложенное выше, конечно, не снимает важности продолжения и, возможно, расширения мониторинга за состоянием экосистем и конкретных видов биологических ресурсов. Существующая система принципов регулирования рыболовства нуждается в развитии и совершенствовании. Например, большим недостатком охранных мероприятий является запрет на лов и выборочная охрана хищников, в результате чего в системе наблюдается дисбаланс. Тем более еще очень далеко до разработки конкретных схем рационального природопользования в морях в целом и до умения управлять биоресурсами. Биоресурсный блок в этих схемах должен включать в оптимальных сочетаниях рациональное рыболовство, аквакультуру и природоохранные усилия.
Содержание |
|